
Установим сигнализация на Газель Некст в Подольске
Установка сигнализации с запуском и турботаймером на автомобиль на Газель Некст. Газель Некст с дизельным двигателем Камминз. Стоит с завода предпусковой подогреватель Вебасто. Центральный замок управляется поворотом ключа в водительской двери. В качестве сигнализации установили StarLine A93. Простая и надёжная система с двусторонней связью, возможностью реализации автозапуска и турботаймера, а также с возможной реализацией управления Webasto по шине W-BUS или по аналоговому входу.
Автозапуск и управление вебасто на Газель
Для того, чтобы подключить систему подогрева к сигнализации, можно воспользоваться дополнительным оборудованием охранной системы, но не все модели СтарЛайн могут поддерживать такую возможность.
Установим на машину сигнализацию Starline A93 с дистанционным автозапуском, таким образом решив проблему холодного салона и непрогретого движка по утрам, садясь в машину на заснеженной стоянке. Наши мастера умеют работать с автосигнализациями, поэтому никаких проблем возникнуть не должно — все сделаем в лучшем виде! А заодно и снимем ролик, где продемонстрируем работу уже установленной системы и некоторые ее особенности.
ГАЗель NEXT автозапуск
Схема для самостоятельной установки сигнализации в автомобиль Газель НЕКСТ 2015 года выпуска. Данная карта монтажа имеет рекомендательный характер, цвета проводов могут отличаться в зависимости от года и комплектации автомобиля.
Блок сигнализации можно установить за приборной панелью, антенну оптимально закрепить на лобовом стекле.
Место установки сигнализации на газель
Под капотом, ниже штатного уплотнителя, под гайку. Снять «пистон» на котором держится жгут проводов, закрепить сирену гайкой, после чего жгут идеально сядет обратно. Концевик капота можно установить в любом удобном месте.
Снять «пистон» на котором держится жгут проводов, закрепить сирену гайкой, после чего жгут идеально сядет обратно. Концевик капота можно установить в любом удобном месте.
Ручной тормоз: розовый/черный провод за приборной панелью, развязать диодом.
Концевики дверей: удобно взять с плафона, или Серый/Белый провод в коричневом разъеме за приборной панелью через резистор 1кОм.
Указатели поворотов: под рулевой колонкой Красный + Желтый +
Центральный замок: в жгуте из двери водителя Желтый/Белый + Желтый/Черный +
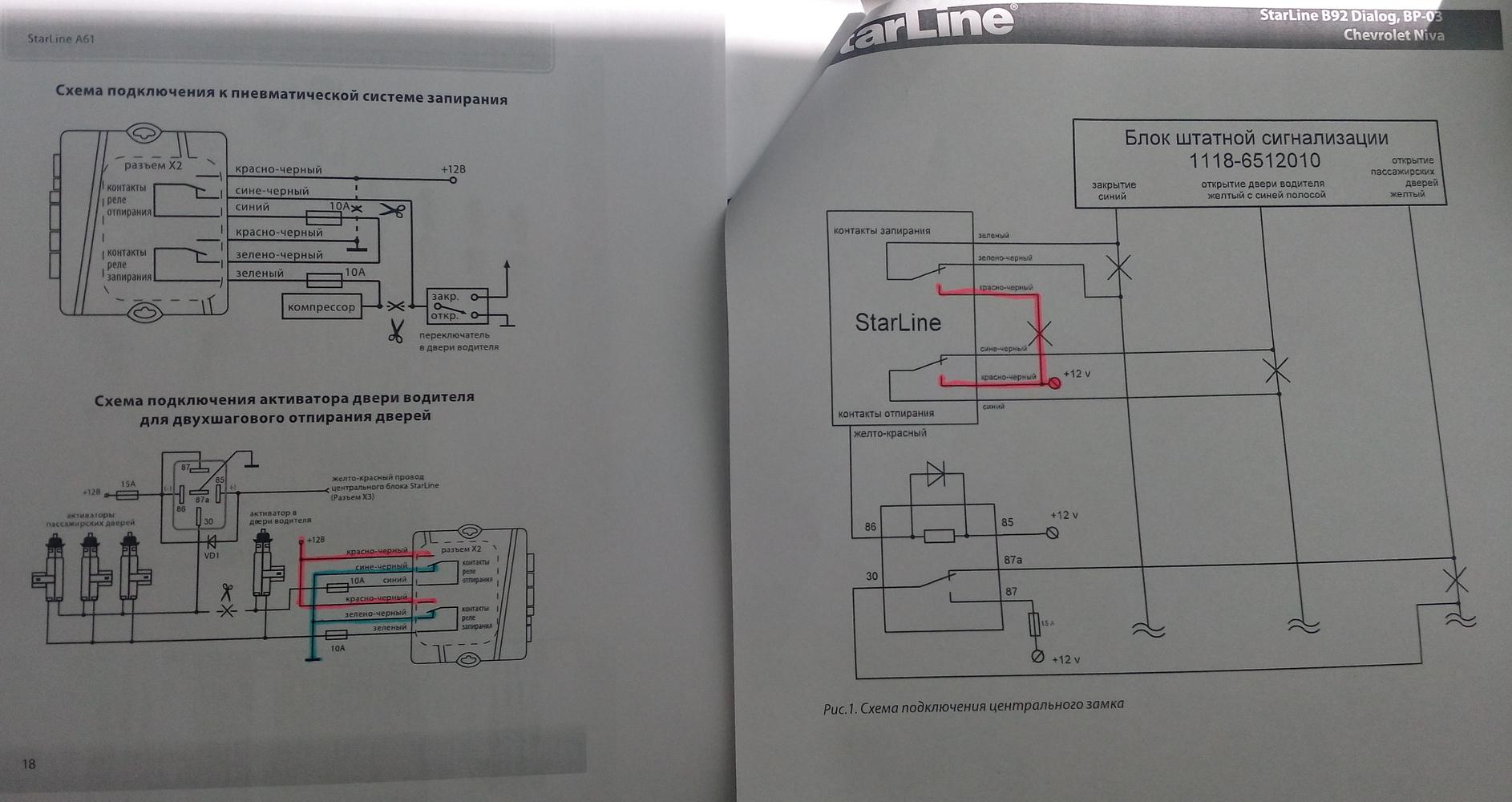
Либо сделать по схеме как на приоре на блоке центрального замка есть Синий/Черный закрыть — Разорвать, открыть — на массу.
Контроль запуска двигателя: под капотом на генераторе, Розовый провод.
Силовые цепи запуска автомобиля: +12В — Желтый Зажигание 1 — Красный Зажигание 2 — Коричневый Стартер — Белый
Сигнализация с установкой на Газель
Точки подключения автосигнализации на Газель NEXT
Замок зажигания:
12V – розовый
IGN1 – красный
IGN2 – коричневый
ST — белый
На приборке:
Повороты — красный и жёлтый/белый
Двери — серый/белый
Ручник — розовый/черный
Центральный замок:
Синий/черный — масса – открывание, разрыв – закрывание
Контроль работы двигателя — желтый, от датчика положения распредвала с задней стороны двигателя.
Установка сигнализации на Газель Некст, точки подключения ГАЗель-Next
Схема установки автосигнализации StarLine B94 на автомобиль ГАЗель-Next с 2012 года
Устанавливаемое оборудование:
1. Снимаем щиток приборов Газель Некст. Для этого демонтируем подторпедник, далее открутив 4 самореза снимаем рулевой кожух. Снимаем накладку щитка приборов и отвернув 4 самореза крепления, снимаем сам щиток
Снимаем накладку щитка приборов и отвернув 4 самореза крепления, снимаем сам щиток
2. Устанавливаем светодиод в левую стойку лобового стекла, сервисную кнопку в любое удобное место, антенну со встроенным датчиком удара и наклона на лобовое стекло
3. Устанавливаем под капотом сирену, концевик капота и датчик температуры двигателя. Провода прокладываем через штатный уплотнитель с левой стороны моторного щита
4. Блок сигнализации устанавливаем за щиток приборов
6. В разъеме щитка приборов подключаем контроль ручного тормоза согласно схеме 1, концевики дверей согласно схеме 2 и аварийную сигнализацию.
7. Контроль работы двигателя подключаем к розовому проводу на генераторе, сигнал «Генератор +»
8. Подключение к штатному центральному замку осуществляем в жгуте идущем из двери водителя. Поодключение делаем согласно схеме 3
9. В жгуте замка зажигания подключаем зажигание, зажигание 2, стартер и питание +12В блока сигнализации и модуля запуска двигателя.
10. Программируем параметры автозапуска (Таблица 2 инструкции по установке):
Программируемая функция Значение функции
№ 09 -длительность прокрутки стартера 6,0 сек
11. Настраиваем датчик удара и наклона. Проверяем работу сигнализации. Сборку салона Газель Некст производим в обратной последовательности.
| Цепь | Цвет | Полярность | Расположение |
| + 12В | бел | Положительная | Замок зажигания |
| стартер | кор | ||
| зажигание | ж | ||
| печка | оранж | ||
| датчик температуры двигателя | серый | красный разъём за панелью приборов | |
| зарядка | розовый | Положительная | черный разъём за панелью приборов |
| давление масла | голубой | ||
| повороты | жёлтый и красный(вместе) | красный разъём за панелью приборов | |
| ручник | голубой | Отрицательная | на таблетке (реле, обеспечивающее мигание лампы STOP) |
| тормоз | зелёный | Положительная | педаль |




Точки подключения автосигнализации на ГАЗ Газель
1. Точки подключения автосигнализации на Газель, Соболь, Баргузин | |||
| Цепь | Цвет | Полярность | Расположение |
| + 12В | бел | Положительная | Замок зажигания |
| стартер | кор | ||
| зажигание | ж | ||
| печка | оранж | ||
| датчик температуры двигателя | серый | красный разъём за панелью приборов | |
| зарядка | розовый | Положительная | черный разъём за панелью приборов |
| давление масла | голубой | красный разъём за панелью приборов | |
| повороты | жёлтый и красный(вместе) | красный разъём за панелью приборов | |
| ручник | голубой | Отрицательная | на таблетке (реле, обеспечивающее мигание лампы STOP) |
| тормоз | зелёный | Положительная | педаль |
2.
 Точки подключения автосигнализации на ГАЗ 3310 Валдай
Точки подключения автосигнализации на ГАЗ 3310 ВалдайЗамок зажигания:
+12 – черный
зажигание 1 – желтый
зажигание 2 – оранжевый
стартер – коричневый
Панель приборов, разъём XP2:
повороты (+) – красный и желтый (пин 2 и 3)
ручник (-) – коричневый (пин 4)
Панель приборов, разъём XP3:
тахометр – красный (пин 5)
Концевики все ставим свои. Привода в двери – по желанию.
3. Карта монтажа автосигнализации StarLine А91 на автомобиль ГАЗ-3302 Газель с двигателем КРАЙСЛЕР
Карта установки автосигнализации StarLine А91 на автомобиль ГАЗ-3302 Газель (с двигателем КРАЙСЛЕР), МКПП, ИММО отсутствует. Для автомобилей с двигателем 406 и более поздних модификаций могут незначительно отличаться цвета проводов, расположение такое же – очень похоже на Волгу-31105.
Для доступа к проводке и внутренним полостям торпедо разбираем салон. Нам понадобятся пластиковая лопатка, отвертка крестовая. Ключ торцовый 7мм.
3.1. Разбираем салон
Начинаем с дверей на примере водительской двери при условии, что будут выполняться работы по установке электроприводов центрального замка.
Фото 1.1. Нужно демонтировать элементы, обведенные овалами. Саморезы по стрелочкам – не выкручивать. В прямоугольнике – нестандартные элементы управления ЭСП (В связи с этим ручки стеклоподъемника нет).
Фото 1.2. Выкручиваем винт М6 шлицевой отверткой и снимаем накладку ручки открытия двери.
Фото 1.3. Выкручиваем два болта М6 (головка 7мм) крепления ручки двери. Снимаем её.
Фото 1.4. Снимаем ручку стеклоподъемника (если есть). После этого сдергиваем обшивку – клипсы по периметру.
Фото 1.5. Выкручиваем саморезы крепления накладки левой стойки лобового стекла и снимаем её.
Фото 1.6. Выкручиваем два болта М6 (головка 7мм) крепления рукоятки открытия капота.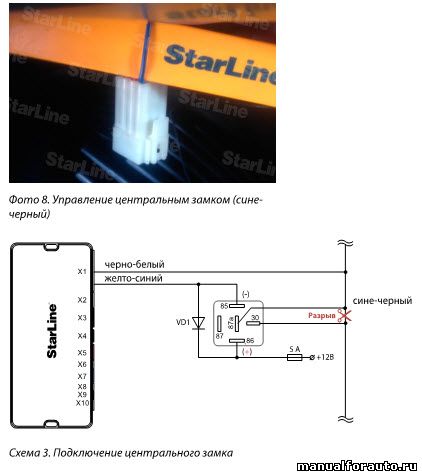 Проверяем – не вкручены ли саморезы крепления акустической системы насквозь в металл.
Проверяем – не вкручены ли саморезы крепления акустической системы насквозь в металл.
Фото 1.7. Выкручиваем саморез крепления накладки левой кик-панели и аналогично – правой над аркой колеса.
Фото 1.8. Выкручиваем еще по одному саморезу крепления накладок над порожком (на примере левой). Снимаем накладки кик-панелей.
Фото 1.9. Извлекаем две заглушки из обрамления под доской приборов.
Фото 1.10. Выкручиваем четыре самореза крепления обрамления доски приборов и извлекаем обрамление.
Фото 1.11. Выкручиваем четыре самореза (головка 7мм) крепления доски приборов и извлекаем её.
Фото 1.12. Выкручиваем два самореза крепления обшивок рулевой колонки и снимаем нижнюю половинку обшивок.
3.2. Приступаем к установке автосигнализации на Газель
Фото 2.1. Просверлив и загрунтовав отверстия 4,2 мм в указанной зоне (помним про лицевую панель двери!), крепим корпус электропривода SATURN, соединяем его тягу с тягой замка и регулируем ход тяг при защёлкнутом механизме замка (как будто дверь захлопнута).
Фото 2.2. Провода из двери в салон протягиваем через штатную проходку. При её отсутствии сверлим отверстия. В зоне перехода из двери в кузов защищаем провод куском пластиковой гофрированной трубки 10мм. Подключение делаем по схеме 2.1.
Провода прокладываем не внатяг (как это сделано на фото – осталось после предыдущей установки). Возможно, что механизм замка придется разбирать, подтачивать и смазывать для обеспечения плавности и легкости движения механизма, а затем и регулировать его после установки в дверь.
Схема 2.1. Управление центральным замком.
Фото 2.3. Концевик AU-29 (PS-8) контроля двери правильнее и проще установить в отмеченной зоне, просверлив отверстие 8,2 мм из салона. Предварительно установите маячок и проверьте – не попадает-ли на концевик отверстия в петле двери. Откорректируйте точное место сверления.
Фото 2. 4. На данном автомобиле уже имелись кнопки типа PS-9 в верхних уголках проема двери. Тянуть до них проводку – непросто.
4. На данном автомобиле уже имелись кнопки типа PS-9 в верхних уголках проема двери. Тянуть до них проводку – непросто.
Фото 2.5. На проводах разъемов приборной доски (вид сзади) подключаем световую сигнализацию, контроль работы двигателя и контроль стояночного тормоза (по схеме 2.2).
Схема 2.2. Контроль стояночного тормоза (ручника).
Фото 2.6. Расстыковываем разъем замка зажигания, поднимаем его в проем приборной доски и подключаем на его пучке силовые провода сигнализации.
Фото 2.7. Заблокировать можно бензонасос. Его провод проходит через разъем под капотом над левым подрамником (под гидроцилиндром сцепления).
Фото 2.8. Провода к сирене, кнопке и датчику температуры в подкапотное пространство можно вывести через проходку над аккумулятором.
Фото 2.9. Еще одно место для вывода проводов под капот – за расширительным бачком — рядом с педальным узлом (как минимум сюда можно вывести провода реле блокировки).
Фото 2.10. Расположение сирены и концевика капота при первом варианте вывода проводов под капот.
Фото 2.11. На площадке электропривода «дворников» можно установить сирену при использовании проходки фото 2.9 (второй вариант).
Фото 2.12. В этом случае концевик можно установить по аналогии с фото 2.10 в имеющееся отверстие. Но правильнее просверлить отверстие рядом, предварительно определив маячком наиболее оптимальное место.
Фото 2.13. Температурный датчик сигнализации крепим или на ГБЦ под один из имеющихся крепежей М6 (как на фото) или на один из патрубков системы охлаждения капроновыми хомутами-стяжками.
Фото 2.14. Датчик удара удачно разместился на трубе рулевой колонки впритык к подрулевым переключателям – здесь он не будет мешать собрать обшивки колонки.
Фото 2.15. Светодиод устанавливаем в накладке левой стойки лобового стекла.
Фото 2.16. Модуль приемо-передатчика наклеиваем на лобовое стекло. Для облегчения прокладки проводов за обивкой потолка ослабляем винты крепления опор противосолнечных козырьков.
Для облегчения прокладки проводов за обивкой потолка ослабляем винты крепления опор противосолнечных козырьков.
Если автомобиль с будкой (термобудкой), то вход сигнализации «контроль багажника» можно использовать для контроля вскрытия будки, установив концевик на дверь будки, распахивающуюся первой.
На автомобилях автобусной компоновки (пассажирские, грузовые и грузо-пассажирские фургоны) со сдвижной и задними распашными дверями нужно будет:
а) Для обеспечения работы электропривода ЦЗ кроме установки ЦЗ в сдвижную дверь использовать специальный подпружиненный сдвоенный контакт в проеме этой двери.
б) Установить концевики в проемы сдвижной и задней двери, которая открывается первой и, разумеется – электропривод ЦЗ в эту дверь.
в) Заменить предохранители на проводах управления ЦЗ с 10А на 15А (четыре электропривода !).
4. Точки подключения автосигнализации на ГАЗель NEXT с дизелем Cummins
Замок зажигания после разъёма:
+12 — Красный
IGN1 — Жёлтый
IGN2 — Оранжевый/чёрный
Starter — Коричневый
Центральный замок:
на блоке центрального замка за рулевой колонкой — синий с чёрной полосой: закрыть — разрыв, открыть — на массу.
Панель приборов:
Генератор — розовый
Ручник — розовый/чёрный, необходимо отвязаться диодом от панели приборов.
Двери общий: серый/белый, необходимо развязаться двумя диодами (один в разрыв от панели приборов, второй катодом к катоду первого диода, анодом к сигнализации). Также вход дверей сигнализации необходимо подтянуть к +12В через резистор примерно в 10кОм, иначе через 10сек после закрытия дверей на этом проводе будет 2,5В и сигнализация будет считать дверь открытой.
5. Точки подключения автосигнализации на Газель NEXT 2014
Замок зажигания:
12V – розовый
IGN1 – красный
IGN2 – коричневый
ST — белый
На приборке:
Повороты — красный и жёлтый/белый
Двери — серый/белый
Ручник — розовый/черный
Центральный замок:
Синий/черный — масса – открывание, разрыв – закрывание
Контроль работы двигателя — желтый, от датчика положения распредвала с задней стороны двигателя.
Установка Сигнализации Газель Газон Некст ⋆ SVS
Наши мастера установят выбранную Вами сигнализацию на Ваш автомобиль Газель, Газон.
Установка Сигнализации на Ваш автомобиль.
Установка сигнализации CENTURION S9 9500₽
(Брелок ж\к. Обратная связь. Турбо-таймер Автозапуск. )
Цена по записи на выходные дни.
Сигнализация с односторонней связью
Это одна из самых первых и в тоже время самых распространенных видов сигнализаций. Хоть данный вид и устарел в плане охраны транспорта, но все равно пользуется спросом у потребителя за счет своей небольшой цены. Функции данного вида сигнализации: звуковые и световые сигналы при попытке взлома дверей, капота, багажника; блокировка двигателя; возможность подключения к центральному замку (открыть, закрыть) машину. В односторонней системе защиты от угона сигнал передается только от брелока к автомобилю. Обратного сообщения нет.
| Модель | Функции | цена |
| CETURION A-95* | Обратной связи нет | 3300₽ |
| Pantera QX-44 | Обратной связи нет | 6500₽ |
| ALLIGATOR A-2s | Обратной связи нет | 6500₽ |
| ALLIGATOR A-1s | Обратной связи нет | 6500₽ |
CETURION A-95 Управление центральным замком с брелока
Сигнализация с обратной связью
Двухсторонняя сигнализация содержит модуль обратной связи. У владельца находится брелок-пейджер, на которые приходит вся информация о текущем состоянии транспорта. Такая система сложнее и дороже, но намного надежней. Важной характеристикой двухсторонней автосигнализации является дальность оповещения и управления. Благодаря этому параметру владелец будет знать, что происходит с его транспортом на достаточно удаленном расстоянии. На жидкокристаллическом экране изображен автомобиль и указан элемент, на который совершено постороннее воздействие. Starline GSM-Управление с телефона.
Starline GSM-Управление с телефона.
| Модель | Функции | Цена |
| Starline A61 | Турбо-таймер | 10500₽ |
| Starline A63 | Турбо-таймер | 11500₽ |
| Starline A63 gsm | Турбо-таймер | 16200₽ |
| Starline E63 | Турбо-таймер | 13300₽ |
| Starline E63 gsm | Турбо-таймер | 18000₽ |
| Tomahawk 7.2 | Турбо-таймер | 10500₽ |
| KGB EX-7 | Обратная связь | 9500₽ |
| CETURION Х-6 | Обратная связь | 8700₽ |
Сигнализация с обратной связью и автозапуском
Двухсторонняя сигнализация с автозапуском содержит модуль обратной связи. У владельца находится брелок-пейджер, на которые приходит вся информация о текущем состоянии транспорта. На жидкокристаллическом экране изображен автомобиль и указан элемент, на который совершено постороннее воздействие. Можно, не выходя из дома запустить мотор, прогреть машину. Snarline GSM-Управление с телефона.
| Модель | Функции | Цена |
| Centurion S-9 | Турбо-таймер Автозапуск | 10500₽ |
| Centurion is-10 | Турбо-таймер Автозапуск | 11500₽ |
| Starline A93 | Турбо-таймер Автозапуск | 12500₽ |
| Starline A93 gsm | Турбо-таймер Автозапуск | 17200₽ |
| Starline E93 | Турбо-таймер Автозапуск | 14100₽ |
| Starline E93 gsm | Турбо-таймер Автозапуск | 18800₽ |
| Tomahawk 9.9 | Турбо-таймер Автозапуск | 11500₽ |
Tomahawk 9. 7 7 | Турбо-таймер Автозапуск | 11500₽ |
| Tomahawk 1031 | Турбо-таймер Автозапуск | 11700₽ |
Подключение управлением Webasto от сигнализации StarLine
Установка на Газель Next ЦМФ.
| Модель | Ц.З три двери | Ц.З. четыре двери |
| 3 места | +3000 ₽ | +5000 ₽ |
| 7 мест | стандарт | +3000 ₽ |
| Фермер | стандарт | -0- |
Может вас заинтересует как экономить топливо. Читать дальше ДА Нет.
| Цепь | Цвет | Полярность | Расположение |
| + 12В | бел | Положительная | Замок зажигания |
| стартер | кор | ||
| зажигание | ж | ||
| печка | оранж | ||
| датчик температуры двигателя | серый | красный разъём за панелью приборов | |
| зарядка | розовый | Положительная | черный разъём за панелью приборов |
| давление масла | голубой | красный разъём за панелью приборов | |
| повороты | жёлтый и красный(вместе) | красный разъём за панелью приборов | |
| ручник | голубой | Отрицательная | на таблетке (реле, обеспечивающее мигание лампы STOP) |
| тормоз | зелёный | Положительная | педаль |
 Как работать с этой машиной, знают многие, а вот как правильно сохранить ее – меньшинство. Обезопасить центральный или любой другой замок может обычная автомобильная сигнализация. Как ее выбрать и подключить, читайте в данной статье.
Как работать с этой машиной, знают многие, а вот как правильно сохранить ее – меньшинство. Обезопасить центральный или любой другой замок может обычная автомобильная сигнализация. Как ее выбрать и подключить, читайте в данной статье. Привода в двери – по желанию.
Привода в двери – по желанию.
 Его провод проходит через разъем под капотом над левым подрамником (под гидроцилиндром сцепления).
Его провод проходит через разъем под капотом над левым подрамником (под гидроцилиндром сцепления). б) Установить концевики в проемы сдвижной и задней двери, которая открывается первой и, разумеется – электропривод ЦЗ в эту дверь. в) Заменить предохранители на проводах управления ЦЗ с 10А на 15А (четыре электропривода !).
б) Установить концевики в проемы сдвижной и задней двери, которая открывается первой и, разумеется – электропривод ЦЗ в эту дверь. в) Заменить предохранители на проводах управления ЦЗ с 10А на 15А (четыре электропривода !).

Наиболее распространенные причины обращения в сервис владельцев автомобиля ГАЗель NEXT.
Наиболее распространенные причины обращения в сервис владельцев автомобиля ГАЗель NEXT.
Ремонт Газели всегда оказывается некстати – приходится останавливать работу, сниматься с рейсов или отменять заказы. Неплохо, если неполадки несерьезные могут быть устранены своими силами, но так бывает далеко не всегда. Самостоятельно отремонтировать коробку передач, двигатель или задний мост с заменого редуктора окажется непросто даже для опытных водителей, однако, существует ряд возможностей по разрешению проблемы.
Неполадки в работе личного автомобиля доставляют массу неудобств, но едва ли могут приводить к таким неприятностям, которые приносит с собой поломка рабочего или коммерческого транспорта.
Существует 3 наиболее распространенные причины обращения владельцев Газель Некст за помощью специалистов станций технического обслуживания и ремонта автотранспорта:
1. Ремонт редуктора заднего или переднего автомобильного моста.
2. Устранение неполадок двигателя внутреннего сгорания автомобиля.
3. Дефектовка и комплексный ремонт коробки переключения передач.
Обращаем ваше внимание, что данные проблемы устранить без помощи специалистов окажется практически невозможно. И дело здесь не только в опыте работы, необходимом для ликвидации поломок этого типа, но и в специализированном оборудовании сервисного центра, используемом мастерами при проведении работ.
Ремонт двигателя Газель
Модельный ряд автомобилей Газель Next оборудован дизельными двигателями Cummins ISF 2.8 массой более 200 кг. Наличие такого агрегата позволяет надолго забыть о проблемах в моторном отсеке, однако, время и нагрузки дают о себе знать. Несвоевременная замена моторного масла или топливного фильтра и кустарное дизельное топливо за непродолжительное время приведут даже новый мотор в непригодное для эффективной работы состояние.
О появлении проблем в работе Каминса свидетельствует появление одного или нескольких типичных симптомов несправного двигателя:
1. Автомобиль перестал заводиться или заводится с перебоями.
2. Двигатель колотит, доносятся непривычные шумы со стороны ДВС.
3. Появление сажевой взвеси в выхлопах авто.
4. Рабочая мощность мотора непривычно мала, двигатель не выходит на норму.
5. Двигатель работает нестабильно, обороты плавают.
Многие из вышеозначенных симптомов могут проявиться сразу после заливки некачественных ГСМ. Поддельное моторное масло не образует прочной защитной пленки на рабочих поверхностях агрегатов ДВС, а отсутствие качественных присадок приводит к закипанию масла или его возгоранию с последующей детонацией топливной смеси. Все это ведет к разрушению турбины Holset HE211W, чья неисправность является одной из наиболее распространенных причин поломки Cummins ISF 2.8.
Если поменять турбину своими руками сможет опытный водитель, то на самостоятельное проведение капитального ремонта двигателя рассчитывать не стоит. Он может потребоваться, когда транспортное средство проехало весь рейс на некачественном моторном масле, а в кузове все это время находился груз с превышением допустимой массы или близкий к ее пределу. Подобная эксплуатация нередко оказывается причиной «боя» цилиндропоршневой группы. Ее восстановление потребует расточки с комплексным капитальным ремонтом.
Если качество ГСМ не вызывает сомнений, то появление стука может быть вызвано неисправными форсунками. Их поэтапное отключение через электронный управляющий модуль позволит установить наверняка, являются ли они причиной «боя». Поменять форсунки самостоятельно получится только при наличии профессионально оборудованного ремонтного бокса или гаража. Аналогичные условия потребуется соблюсти и для замены неисправного топливного насоса или рампы.
Их поэтапное отключение через электронный управляющий модуль позволит установить наверняка, являются ли они причиной «боя». Поменять форсунки самостоятельно получится только при наличии профессионально оборудованного ремонтного бокса или гаража. Аналогичные условия потребуется соблюсти и для замены неисправного топливного насоса или рампы.
Ремонт КПП Газель
В автомобилях Газель Next устанавливается пятиступенчатая механическая коробка переключения передач. Если обратить внимание на цены МКПП, то нетрудно заметить, что данная деталь относится к числу наиболее дорогостоящих агрегатов транспортного средства. Утечки трансмиссионного масла из системы или несвоевременная его замена – одни из основных причин возникновения неисправностей коробки. Другим немаловажным факторов, приводящим к поломке КПП является агрессивное вождения авто и приложение чрезмерных усилий к рычагам устройства.
Симптомами неисправностей МКПП являются выпадающие скорости, возникновение проблем с их переключением и посторонние звуки (дребезжание или звон), доносящиеся со стороны коробки. Проведение самостоятельного ремонта возможно, если речь идет о смене внешних компонентов передаточной системы. Когда дефекты затрагивают внутренние элементы КПП, тогда необходим полный разбор устройства с последующей дефектовкой и выявлением изношенных узлов. Такая процедура выполняется только мастерами автомеханиками на СТО.
Если вы заметили у своего автомобиля признаки неисправности по данным проблемам, следует незамедлительно обратится в сервисный центр для диагностики и ремонта!
Напоминаем, что в нашем дилерском центре действует скидка на постгарантийный период 15%!
Как установить будильник Apple Watch за 3 простых шага
Изящный дизайн и универсальность Apple Watch вызывают трепет у многих. Устройство позволяет общаться без помощи рук и выполнять любую задачу, что идеально подходит для предпринимателя в дороге! Однако даже этому занятому бизнес-магнату время от времени нужно спать. К счастью, в Apple Watch есть функция будильника Apple Watch.
К счастью, в Apple Watch есть функция будильника Apple Watch.
Apple Watch можно использовать как будильник на прикроватной тумбочке, пока они заряжаются на боку.Часы оснащены ненавязчивыми часами, в которых, в отличие от мобильного телефона, вам не нужно брать трубку или нажимать кнопку, чтобы увидеть время — просто подтолкните часы, чтобы проверить время, затем переверните и получите еще несколько Z!
Установка будильника Apple Watch — это всего лишь три простых шага!
- Установите часы в ночной режим
Положив часы на бок на прикроватной тумбочке, подключите Apple Watch к зарядному устройству. После начала зарядки часы автоматически перейдут в ночной режим.Дисплей отключается, когда он не используется, но его можно снова включить, если его коснуться или даже просто подтолкнуть. - Установить будильник
Вам нужно будет установить или настроить будильник после того, как часы перейдут в ночной режим. Сначала откройте приложение «Будильник» на Apple Watch, затем с усилием нажмите на панель дисплея и нажмите «добавить будильник». Либо поверните колесико Digital Crown, расположенное сбоку на часах, либо коснитесь экрана, чтобы настроить будильник.
Вы также можете удерживать цифровую корону, пока не появится Сири, и попросить ее установить для вас будильник.
Presto! Тебе хорошо спать! - Отложить
Хотите поймать несколько лишних Z по утрам? Перед включением будильника дисплей станет ярче, а текст станет желтым. Затем Apple Watch издадут звуковой сигнал. Чтобы отложить Apple Watch на девять минут, нажмите колесико Digital Crown.
Чтобы выключить будильник, нажмите боковую кнопку.
Настроить будильник Apple Watch так же просто, как 1-2-3. Просто установите, спите и отложите (если позволяет ваше время.) Или подпрыгните и начните свой день!
Какое руководство от Apple вы хотели бы увидеть в следующий раз? Есть ли какие-нибудь советы или уловки, о которых вы хотите узнать? Дайте нам знать!
Фотография предоставлена Flickr
Комментарии
комментария
ГАЗ: в Германии поступят в продажу электрические фургоны ГАЗель NEXT
EFA-S GmbH, немецкий партнер ГАЗа, строит электромобили на базе российского LCV ГАЗель NeXT производства Горьковского автозавода. В дополнение к базовой версии E35 с откидным бортом и изолированному фургону с брезентовым покрытием E35, представленной EFA-S прошлой осенью, теперь в Германии доступны цельнометаллические фургоны, автобусы-фургоны до 9 мест и версия с брезентовым верхом с двойной кабиной. .
В дополнение к базовой версии E35 с откидным бортом и изолированному фургону с брезентовым покрытием E35, представленной EFA-S прошлой осенью, теперь в Германии доступны цельнометаллические фургоны, автобусы-фургоны до 9 мест и версия с брезентовым верхом с двойной кабиной. .
EFA-S GmbH пополнила семейство электромобилей на базе ГАЗель NEXT тремя новыми членами, их полезная нагрузка увеличилась почти вдвое: максимальная полная масса всех автомобилей E35 GAZelle NEXT составляет 3,5 т или 4,25 т, что увеличивает их фактическую грузоподъемность на 750. кг.
Леонид Долгов, директор ГАЗ Интернешнл:
— Благодаря партнерству с EFA-S мы делаем большой прыжок в будущее, совместными усилиями создаем абсолютно бесшумные, экологичные и экономичные коммерческие автомобили на базе наших новая ГАЗель NEXT семейства.Осенью прошлого года EFA-S начала продажи бортовых электромобилей E35 на базе ГАЗели NEXT, а сегодня у нас появились новые члены в этом семействе, и мы первые на немецком рынке коммерческих автомобилей, предложившие 7-местный электромобиль. Кроме того, мы являемся единственным проектом в Германии по производству электромобилей, что означает, что мы можем предложить все виды надстроек, которые нужно добавить, создавая индивидуальный коммерческий автомобиль, отвечающий всем потребностям наших бизнес-клиентов.
Теперь, когда в линейке электрических устройств появилось больше моделей, EFA-S может предлагать индивидуальные решения, отвечающие конкретным бизнес-потребностям своих клиентов.
Бастиан Бойтель, генеральный директор EFA-S:
— Все базовые версии электромобилей E35 ГАЗель NEXT имеют грузоподъемность около 1 тонны и могут проехать 200 км на одной зарядке, чего достаточно для большинства задач. Для этой цели EFA-S использует литий-железо-фосфатные батареи нового поколения мощностью 80 кВтч с керамическими сепараторами (LFP). По желанию заказчика EFA-S установит батареи еще большего размера, до 244 кВтч. EFA-s предлагает бесшумные экологически чистые и экономичные электромобили на базе ГАЗель NEXT.
действительно безопасны и не требуют никеля и кобальта, которые часто вызывают некоторые проблемы. С новым аккумулятором EFA-s предлагает вдвое большую мощность аккумулятора, чем когда-либо ранее был стандартным для электрических фургонов.
ИСТОЧНИК: ГАЗ
Газелей смотрят друг другу в глаза, чтобы сохранить статус-кво — и мы делаем то же самое — Quartz
Мы можем представить, что мы независимые мыслители, но когда дело доходит до столкновения, мы часто заканчиваем тем, что идем вместе с толпой .Это потому, что, как и у газелей, у нас есть биологическая мотивация держаться за свои стада.
Газель придерживается своего вида, как будто от нее зависит ее жизнь — потому что с точки зрения генов это так. У некоторых газелей на заднице черные полосы, у других — белые, а у некоторых — одна черная и одна белая. Эти полосы показывают экологическую нишу, к которой приспособлена газель, а спаривание с генами для правильной ниши создает более сильное потомство. Но газели не думают о зачатии концептуально — они просто сканируют окурки.
Газель всегда хочет, чтобы ее окружали правильные друзья. Если он уходит сам по себе, возникает чувство тревоги, и кортизол, химическое вещество стресса, резко возрастает. Повторное соединение с газелями, похожими на задницу, снимает этот дискомфорт с помощью удобного окситоцина: они прикрывают вашу спину, потому что безопасность в количестве.
Газели всю жизнь продвигаются к центру стада, где безопаснее. Когда они слишком стары, чтобы толкаться, они оказываются за границами и встречают свою судьбу. Чтобы не остаться позади, газели также убегают от хищников, когда их товарищи встревожены.Некоторые существа когда-то сопротивлялись этому, отказываясь бежать, если только они не обнаружили угрозу собственными чувствами. Но этих тварей часто застали врасплох — и съели заживо. Таким образом, у млекопитающих развился мозг, который доверяет сигналам тревоги своих товарищей.
Хватит! , возможно, вы захотите сказать. Я хочу исследовать пышные луга, а не следовать за стаей. Но каждый раз, когда вы самостоятельно отклоняетесь от курса, вас пронзают угрожающие чувства, которые невозможно объяснить.
Казалось бы, есть несколько преимуществ в том, чтобы идти в одиночку к более зеленым пастбищам вместо того, чтобы держаться за стадо. Таким образом, газель сможет спокойно есть свежую траву вместо того, чтобы соревноваться за ту же самую траву, которую уже вытаптывали ее товарищи по стаду.Но когда он отдаляется от своего стада, его окситоцин падает, и его тревога по кортизолу срабатывает.
В естественном состоянии изолированное млекопитающее живет недолго. Естественный отбор создал мозг, который пугает вас опасными химическими веществами всякий раз, когда вы уходите, и награждает вас большим потоком окситоцина, когда вы возвращаетесь. Эта нейрохимическая операционная система побуждает млекопитающих искать спасения в числах.
Газели — млекопитающие, и вы тоже млекопитающее. Вашими химическими веществами управляют структуры мозга, общие для всех млекопитающих, но вы ненавидите эти нейрохимические американские горки.Так что же делать большому мозгу млекопитающему?
Ваш мозг любит искать подходящие факты, потому что это чувство принадлежности стимулирует ваш окситоцин; он находит доказательства угрозы, потому что это снижает уровень кортизола. Это работа, для которой эволюционировала ваша кора головного мозга. Возможно, вы сами искали социальной поддержки только для того, чтобы обнаружить, что вашу задницу осуждают. (И, по правде говоря, вы тоже часто проверяете полосы на чужих задницах.)
Как человек, когда вы видите, как другое стадо топчет, вы, вероятно, думаете, что они дураки; вы можете даже подумать, что их лидеры придумывают угрозы, чтобы удержать последователей в своей лоне.Но когда ваше стадо в панике, это ощущается по-другому.
Газели не питают иллюзий по поводу того, каким должно быть стадо. Они не растут, наблюдая за Друзьями и ожидая, что другие будут рядом с ними. Возможно, вам не удастся избежать роли, которую ваша кора головного мозга эволюционировала, но вы можете управлять своим стадным менталитетом с ловкостью прыгающей газели.
Они не растут, наблюдая за Друзьями и ожидая, что другие будут рядом с ними. Возможно, вам не удастся избежать роли, которую ваша кора головного мозга эволюционировала, но вы можете управлять своим стадным менталитетом с ловкостью прыгающей газели.
эволюция голосовой тревожной связи у грызунов | Behavioral Ecology
Почему животные издают потенциально опасные сигналы тревоги, на протяжении десятилетий озадачивала эволюционных биологов (Klump and Shalter, 1984; Maynard Smith, 1965), и понимание адаптивной полезности тревожной коммуникации оказало влияние на объяснение эволюции социального поведения через родство. отбор (Келлер и Рив, 2002).Начиная с классических исследований Шермана (1977) и Данфорда (1977), которые продемонстрировали кумовские преимущества от вызова сусликов, другие обнаружили доказательства того, что животные получают как прямую, так и косвенную приспособленность (Браун, 1987), испуская потенциально опасные звуковые сигналы тревоги (Блюмштейн и др. , 1997; Hoogland, 1995; Schwagmeyer, 1980). Поэтому удивительно, что эволюционное происхождение сигнала тревоги осталось практически неизученным.
Методы изучения текущей адаптивной полезности теоретически просты и включают корреляционные и экспериментальные компоненты: (1) выдвинуть гипотезу об адаптивной функции, (2) найти корреляцию между вариациями в выражении признака и эволюционной приспособленностью и (3) манипулировать признаком. экспрессия для демонстрации причинно-следственной связи (Tinbergen, 1963; Tinbergen et al., 1962). Однако демонстрация начальных условий, благоприятствующих развитию признака, явно коррелятивна. Вопросы эволюции, подобные этим, изучаются с помощью сравнительного метода, в котором выявляется распределение признаков среди многих видов и формально проверяются гипотезы о коэволюции (Brooks, McLennan, 1991; Harvey and Pagel, 1991; Maddison and Maddison, 2001). ). Эволюция носит кумулятивный характер, и условия, благоприятствующие начальному развитию признака и его последующему поддержанию, не обязательно должны быть идентичными (Дарвин, 1859; Рив и Шерман, 1993). Проблема понимания исходной эволюционной функции состоит в том, чтобы создать гипотезы и переменные, подходящие для сравнительного анализа, которые проверяют дополнительные адаптивные гипотезы. Существуют две основные неисключительные гипотезы, объясняющие нынешнюю адаптивную полезность вызова тревоги: общение с хищниками, чтобы препятствовать преследованию (Hasson, 1991), и общение с сородичами, чтобы предупредить их об опасности (Maynard Smith, 1965, Zuberbühler et al., 1999). .
Проблема понимания исходной эволюционной функции состоит в том, чтобы создать гипотезы и переменные, подходящие для сравнительного анализа, которые проверяют дополнительные адаптивные гипотезы. Существуют две основные неисключительные гипотезы, объясняющие нынешнюю адаптивную полезность вызова тревоги: общение с хищниками, чтобы препятствовать преследованию (Hasson, 1991), и общение с сородичами, чтобы предупредить их об опасности (Maynard Smith, 1965, Zuberbühler et al., 1999). .
Имеются многочисленные свидетельства того, что животные снижают риск для себя, испуская сигналы тревоги (Blumstein, 1999; Hasson, 1991) и, при определенных обстоятельствах, не производят сигналов тревоги, потому что, поступая так, они сделают себя еще более уязвимыми для хищников (Caro , 1995).Поскольку для успеха охоты часто требуется элемент неожиданности, люди могут подавать сигнал тревоги, чтобы предупредить потенциального хищника о том, что он был обнаружен. Сигналы тревоги могут также передавать хищнику информацию о физической способности вызывающего ускользнуть от захвата или защитить себя (Fitzgibbon and Fanshawe, 1988). Сдерживая нападение хищника, звонящий получает личную пользу в фитнесе. Если мы предположим, что тревожный вызов эволюционировал для передачи информации хищникам, мы ожидаем, что вызывающие люди будут пытаться минимизировать свой риск при вызове.Один из способов сделать это — петь только тогда, когда есть возможность визуально определять местонахождение и отслеживать хищников, поскольку визуальное обнаружение хищников может более точно оценить риск нападения хищников (Лима, 1988a, b). Поскольку есть данные о нескольких видах, которые визуально оценивают относительный риск нападения хищников и звонят только тогда, когда они не подвергаются чрезмерному риску (Blumstein and Armitage, 1997; Wolff, 1980), мы можем предсказать, что виды, вызывающие тревогу, будут вести дневной образ жизни, и тревожный сигнал бывает редким или отсутствует у ночных видов, которые ограничены в своей способности надежно оценивать риск нападения хищников и управлять им.
Внутренний риск вызова тревоги также может быть компенсирован потенциальной кумовской пользой (Sherman, 1977; 1980). Вместо или в дополнение к общению с хищником, вызывающий может предупредить сородичей об опасности. Если родственники убегают в безопасное место в ответ на звонок, вызывающий получает полную пригодность благодаря их выживанию. Если бы тревожный вызов развился через кумовство, мы бы ожидали увидеть, что социальные виды (особенно живущие рядом с родственниками) будут звонить с большей вероятностью, чем несоциальные виды.
Грызуны представляют собой отличную модельную систему для изучения эволюции сигналов тревоги, поскольку они различаются по степени своей вокальной, дневной и социальной активности. Мы суммировали эти данные для 209 видов грызунов из 24 различных семейств и использовали нефилогенетические и филогенетические тесты, чтобы исследовать связи между эволюцией дневной жизни, социальным образом жизни и тревогой.
Для выборки всех грызунов, включая тех, которые, как известно, вызывают тревогу, и тех, которые нет, учетные записи видов с номерами от 1 до 702 в видов млекопитающих (ссылки доступны в Приложении, а Млекопитающие неотропических тропических лесов (Emmons, 1997) были осмотрел.Мы определили сигнал тревоги как шум, обычно громкий, издаваемый при обнаружении хищника. Для большинства из 209 видов, включенных в наш окончательный анализ, в наших справочных материалах указано, что животные издают сигналы тревоги в присутствии хищников. Когда у нас были хорошие описания звуков, издаваемых конкретным видом в различных контекстах, и не было упоминания о тревожных сигналах, или когда в источниках конкретно указывалось, что этот вид не известен как тревожный сигнал, мы классифицировали его как не тревожный. Когда у нас было мало или неоднозначных данных о звуках, издаваемых видом, мы считали их неизвестными и удаляли их из набора данных.«Крики страха» или другие защитные звуки (например, стук зубов) не считались сигналом тревоги. Виды, для которых мы не были уверены в контексте, или те виды, которые, по описаниям, издают звуки, когда их держит в руках человек, не были отнесены к категории вызывающих тревогу. Были обобщены данные о моделях активности, социальности, вокализациях и контексте, в котором были изданы вокализации. В различных руководствах и обзорах (ссылки доступны в Приложении) описаны дополнительные виды грызунов, которые издают сигналы тревоги, а также данные для заполнения пробелов в отношении моделей активности, общительности и вокализации.Были проведены консультации со специалистами (см. «Благодарности») для получения дополнительных данных по нескольким видам.
Виды, для которых мы не были уверены в контексте, или те виды, которые, по описаниям, издают звуки, когда их держит в руках человек, не были отнесены к категории вызывающих тревогу. Были обобщены данные о моделях активности, социальности, вокализациях и контексте, в котором были изданы вокализации. В различных руководствах и обзорах (ссылки доступны в Приложении) описаны дополнительные виды грызунов, которые издают сигналы тревоги, а также данные для заполнения пробелов в отношении моделей активности, общительности и вокализации.Были проведены консультации со специалистами (см. «Благодарности») для получения дополнительных данных по нескольким видам.
Мы провели два дополнительных анализа. Во-первых, для нашего набора данных по 209 видам те, которые, как сообщалось, были активны не менее 50% дня, были засчитаны как дневные; те, которые активны преимущественно ночью, как и ведущие ночной образ жизни. Во-вторых, мы изменили наше определение суточности. Согласно нашему строгому определению, мы классифицируем вид как дневной, если никогда не сообщалось, что он активен в ночное время. Это уменьшило размер нашей выборки до 156 видов.Виды, которые могут быть найдены рядом с родственниками, потому что либо они жили семейными группами, либо они жили колониями, либо собирали пищу в скоплениях, оценивались как социальные. Животные, которые, как сообщалось, были обнаружены только поодиночке или парами, или животные, которые, как сообщалось, были территориальными и неколониальными, были признаны не социальными. Определяя социальность таким образом, мы классифицируем как социальные виды, такие как одиночные, но колониальные, суслики ( Spermophilus spp.), Которые были предметом многих исследований адаптивного значения призвания, и мы классифицируем как несоциальные виды, такие как Ондатры ( Ondatra zibethicus ) и дикобразы Северной и Южной Америки ( Erithizon dorsatum и Coendou spp.), которые неизбежно включают виды, нуждающиеся в некоторой степени материнской заботы.
Непотические выгоды от звонка могли развиваться или сохраняться за счет выгоды от предупреждения уязвимых потомков в период родительской опеки (Dawkins, 1979; Hamilton, 1964a, b). Однако у некоторых белок (Swaisgood et al., 1999) матери направляют антихищническое поведение и крики специально на хищников в то время, когда они заботятся об уязвимом и невосприимчивом потомстве. Виды, о которых сообщалось о тревоге в присутствии хищника или других помехах, оценивались как тревожные звонки, тогда как виды оценивались как не тревожные, если были данные вокализации, но тревожные звонки не сообщались.
Используя филогению, изложенную в McKenna and Bell (1997), которая обновляет филогению Симпсона (1945), мы классифицировали по роду 209 видов, по которым у нас были полные и однозначные данные. Это разрешило большинство родов, которые были представлены одним или двумя видами. Роды с более чем двумя видами остались неразрешенными политомиями; многие из них впоследствии были разрешены с помощью филогенетических гипотез на уровне видов, предложенных в различных исследованиях. Как правило, только одной дополнительной филогении было достаточно для разделения видов в род.Следующие критерии применялись в указанном порядке для разрешения любых политомий на уровне рода: (1) молекулярные гипотезы использовались вместо морфологических гипотез; (2) более поздние гипотезы использовались вместо более поздних гипотез; и (3) при прочих равных условиях приоритет отдавался гипотезам, построенным с использованием методов экономии, причем наиболее желательным был консенсус между одинаково экономными деревьями. Мы также исследовали влияние двух недавних молекулярных филогений голарктических сусликов (Harrison et al., 2003; Herron et al., 2004) добился наших основных результатов, изменив наше дерево там, где это необходимо, и повторно выполнив все анализы.
1. Sciurus : Две молекулярные гипотезы (Oshida and Masuda, 2000), использующие методы экономии и вероятности. Обе гипотезы следуют одной и той же структуре, но метод правдоподобия разрешает политомию четырех видов.
Обе гипотезы следуют одной и той же структуре, но метод правдоподобия разрешает политомию четырех видов.
2. Marmota : две молекулярные гипотезы, использующие экономию (Kruckenhauser et al., 1999), а затем методы правдоподобия (которые дополнительно разрешают политомию трех видов; Steppan et al., 1999).
3. Spermophilus : Молекулярная гипотеза с использованием методов экономии (Kruckenhauser et al., 1999) и составленная гипотеза (Blumstein and Armitage, 1998).
4. Cynomys : Скомпилированная гипотеза (Blumstein and Armitage, 1998).
5. Тамиас : Молекулярная гипотеза с использованием методов строгого консенсуса (Piaggio and Spicer, 2001).
6. Neotoma : две молекулярные гипотезы с использованием методов строгого консенсуса (Hayes and Harrison, 1992) и методов экономии (Shipley et al., 1990).
7. Peromyscus : Молекулярная гипотеза с использованием методов экономии (Hogan et al., 1997).
8. Reithrodontomys : две молекулярные гипотезы с использованием методов экономии (Bell et al., 2001) и фенограммы (Nelson et al., 1984).
9. Microtus : две молекулярные гипотезы с использованием методов консенсуса и правдоподобия (Conroy and Cook, 2000). Обе гипотезы следуют одной и той же структуре, но методы правдоподобия разрешают большую политомию.
10. Gerbillurus : Молекулярная гипотеза с использованием методов экономии (Qumsiyeh et al., 1991).
11. Rattus : Молекулярная гипотеза с использованием методов UPGMA (Baverstock et al., 1986).
12. Notomys : Морфологическая гипотеза с использованием методов экономии (Watts et al., 1992).
13. Geomys : две молекулярные гипотезы с использованием методов консенсуса и объединения соседей (Jolley et al. , 2000). Обе гипотезы следуют одной и той же структуре, но методы соединения-соединения разрешают большую политомию.
, 2000). Обе гипотезы следуют одной и той же структуре, но методы соединения-соединения разрешают большую политомию.
14. Pappogeomys : перерисованная морфологическая гипотеза (DeWalt et al., 1993; Russell, 1968).
15. Perognathus : Молекулярная гипотеза с использованием методов экономии (Риддл, 1995).
16. Chaetodipus : Молекулярная гипотеза с использованием методов правдоподобия (Riddle et al., 2000).
17. Dipodomys : Морфологическая фенограмма. Для разрешения большинства видов в пределах Dipodomys spp . Мы решили использовать морфологическую гипотезу (Carrasco, 2000), а не молекулярную гипотезу (Mantooth et al., 2000) того же года, потому что морфологическое дерево было более полные и включали гораздо больше видов из этого рода. Тем не менее, оба источника согласились с размещением всех видов Dipodomys , кроме одного.
18. Ctenomys : Молекулярная гипотеза с использованием методов экономии (Slamovits et al., 2001).
19. Proechimys : Молекулярная и морфологическая гипотеза, объединяющая два вида отдельно от других (Gardner and Emmons, 1984).
Точные тесты Фишера и логистический регрессионный анализ использовались для определения того, было ли на наличие или отсутствие сигнала тревоги повлияло поведение или социальная принадлежность. P Значения <0,05 считались значимыми, а значения p — от.05 – .10 были незначительно значимыми. Поскольку такой видовой анализ не может определить направленность эволюции признака, и такой анализ потенциально затруднен, потому что он не учитывает филогенетическую независимость между близкородственными видами (Harvey and Pagel, 1991), два филогенетических анализа (проведенных с нашими филогенез и модифицированный на основе данных Herron et al., 2004) также использовались для изучения эволюции сигналов тревоги у грызунов.
Тест концентрированных изменений (Мэддисон, 1990) использовался для определения вероятности того, что эволюция сигнала тревоги была сконцентрирована на тех частях филогенетического дерева, в которых присутствовали дневные виды или виды, которые могут быть найдены рядом с родственниками.Тест требует полностью разрешенной филогении, поэтому для разрешения любых оставшихся политомий использовалась опция случайного разрешения в MacClade версии 4.03 (Мэддисон и Мэддисон, 2001). Три дихотомических признака были оптимизированы на разрешенном дереве, чтобы восстановить состояния предков для каждого. В некоторых случаях строгая экономия не могла полностью разрешить реконструкции. Таким образом, алгоритмы ACCTRAN и DELTRAN были применены к каждому дереву символов, что привело к шести реконструкциям состояния предка.Алгоритм ACCTRAN ускоряет изменения характеристик по направлению к корню дерева, максимизируя ранние выгоды и форсируя ранние последующие развороты. Алгоритм DELTRAN, с другой стороны, задерживает изменения в признаках от корня, тем самым максимизируя параллельные изменения (Мэддисон и Мэддисон, 2001).
Большое количество видов препятствовало использованию теста концентрированных изменений для расчета точной вероятности распределения признаков. Вместо этого была использована опция моделирования «фактических изменений» для 10 000 повторов для оценки значений p для каждой реконструкции.Кроме того, чтобы учесть неправильное разрешение состояния предка, моделирование выполнялось с выбранной опцией «либо предка». Чтобы свести к минимуму возможность ложной интерпретации результатов как значимых, консервативный подход поддерживался двумя способами. Во-первых, поправка Бонферрони была применена к критическому значению p для гипотез, проверенных с использованием реконструкций ACCTRAN и DELTRAN (новое значение p , критическое 0,05 / 2 = 0,025). Во-вторых, при вычислении значения p в MacClade использовалось меньше и столько же выигрышей в выделенном характере, а также меньше, чем столько или больше потерь в отличительном характере, чем фактически подсчитано в нашем анализе.
Хотя тест концентрированных изменений позволял сравнивать распределения двух признаков на филогенетическом дереве (Swofford and Maddison, 1987), тест условных состояний (Sillén-Tullberg, 1993) позволял использовать филогенетическую реконструкцию признаков, чтобы спросить, есть ли переход одного символа от нуля к единице или от единицы к нулю или отсутствие перехода с одинаковой вероятностью произойдет в любом состоянии другого символа. Таким образом, это указывало на вероятность того, что эволюционное происхождение данного признака предшествовало эволюции другого признака.Основное предположение состоит в том, что каждая ветвь имеет равную вероятность перехода между состояниями. Чтобы понять направленность эволюции трех признаков, для каждой реконструкции была проведена серия тестов парных условных состояний с использованием CoSta версии 1.03 (Lindenfors, 1999). p Значения меньше, чем 0,025 с поправкой Бонферрони, были интерпретированы как значимые, а p <0,05 как незначительные.
Используя точные тесты Фишера, мы обнаружили значимую связь между дневной жизнью и тревожным звонком, социальной жизнью и тревожным звонком, дневным днем и социальным поведением (Таблица 1).Тем не менее, суточные колебания в зависимости от того, вызвана ли тревога вида, в три-шесть раз больше, чем социальность (Таблица 2). Логистический регрессионный анализ также позволил нам изучить независимое влияние суточного режима и продемонстрировал, что и суточный, и социальный характер могут объяснять значительные различия в том, вызывается ли тревога вида или нет. Однако дневной образ жизни был относительно более важным для объяснения существующей модели тревожных звонков у грызунов (таблица 2), и этот вывод стал еще сильнее, когда мы использовали более строгое определение дневного времени.
В обоих наборах данных (т.е. 209 видов против 156 наборов данных), используя тест концентрированных изменений, мы обнаружили, что реконструкции ACCTRAN ( p <0,001) и DELTRAN ( p <0,0001) показали значительно больше видов, эволюционирующих, вызывая тревогу, вызывая ветви дерева, проявляющие суточный ритм, чем можно было бы ожидать случайно. Тревожный вызов также значительно чаще развивался на ветвях дерева, проявляющих социальность (ACCTRAN, p <.0001; ДЕЛЬТРАН, стр. <0,0001). Кроме того, значительно большее количество социальных видов было сконцентрировано на ветвях дерева, характеризующихся суточной продолжительностью (ACCTRAN, p <0,0001; DELTRAN, p <0,0001).
Тревожный вызов также значительно чаще развивался на ветвях дерева, проявляющих социальность (ACCTRAN, p <.0001; ДЕЛЬТРАН, стр. <0,0001). Кроме того, значительно большее количество социальных видов было сконцентрировано на ветвях дерева, характеризующихся суточной продолжительностью (ACCTRAN, p <0,0001; DELTRAN, p <0,0001).
Тест условных состояний позволил нам определить направленность этого паттерна (таблица 3). В обеих реконструкциях ACCTRAN (рис. 1) и DELTRAN суточность предшествовала эволюции сигнала тревоги. Когда мы использовали реконструкции ACCTRAN, социальность не предшествовала эволюции вызова тревоги.Напротив, реконструкции ДЕЛТРАНА выявили тенденцию к социальности, предшествовавшей эволюции вызова тревоги. Как для ACCTRAN, так и для DELTRAN-реконструкций состояний характера, виды, которые вели дневной образ жизни, не имели большей вероятности эволюционировать в социальную жизнь.
Результаты нашего анализа согласуются с гипотезой о том, что изменение суточного режима предшествовало эволюции сигнала тревоги у грызунов. В целом социальность также оказалась связана с эволюцией вызова тревоги; однако результаты тестов с условными состояниями показали, что виды, которые развили социальность, были, в лучшем случае, лишь незначительно более склонны к развитию вызова тревоги.
Хотя как дневные, так и ночные животные обладают сенсорными способностями, которые хорошо функционируют в соответствующие периоды активности, зрение является ключевым способом точной оценки и динамического отслеживания риска хищничества (Lima, 1988a, b). Поскольку все эти тесты показали, что эволюция суточного режима предшествовала эволюции сигнала тревоги, и поскольку предшествующие доказательства того, что жертва выдает сигнал тревоги только тогда, когда есть достаточно света для обнаружения и отслеживания хищников (Blumstein and Armitage, 1997; Wolff, 1980), мы предлагаем этот сигнал тревоги мог развиться как средство общения с хищниками. Если информация о пригодности вызывающего абонента и его настороженности передается потенциальному хищнику посредством сигнала тревоги, вызывающий, вероятно, препятствует преследованию и увеличивает шансы его выживания (Blumstein, 1999; Cresswell, 1994; Fitzgibbon and Fanshawe, 1988; Хассон, 1991). Такая сигнализация обнаружения может быть исходной функцией тревожной сигнализации у грызунов.
Если информация о пригодности вызывающего абонента и его настороженности передается потенциальному хищнику посредством сигнала тревоги, вызывающий, вероятно, препятствует преследованию и увеличивает шансы его выживания (Blumstein, 1999; Cresswell, 1994; Fitzgibbon and Fanshawe, 1988; Хассон, 1991). Такая сигнализация обнаружения может быть исходной функцией тревожной сигнализации у грызунов.
У некоторых видов сигнал тревоги мог также развиться независимо, чтобы общаться с сородичами, чтобы предупредить их об опасности.В этих случаях неотъемлемый риск вызова тревоги может быть компенсирован потенциальной пользой для семейной пригодности (Dunford, 1977; Sherman, 1977). Наши результаты не могут исключить гипотезу о том, что эволюция призыва для предупреждения уязвимого потомства (см. Blumstein et al., 1997) стала возможной только после того, как повысилась безопасность, связанная с дневным режимом. Однако, если бы основной функцией звонка было предупреждение уязвимого потомства, мы могли бы ожидать, что практически все дневные виды должны вызывать тревогу, потому что у всех в какой-то момент своей жизни есть уязвимое потомство.То, что они этого не делают, говорит нам о том, что что-то еще может быть важным. Кроме того, отцовская забота у млекопитающих не получила широкого распространения, но самцы многих видов вызывают тревогу. Имеющиеся данные не позволяют провести подробный анализ половых различий в тревожных звонках, но наблюдение, которое звонят самцы несоциальных видов (Burke da Silva et al., 1994), может предоставить дополнительные доказательства против необходимости кумовского происхождения звонка.
В рамках этой филогенетической гипотезы мы видим определенные виды, которые развили дневной образ жизни, тревожный вызов и социальность, но впоследствии вернулись к ночному образу жизни, сохраняя при этом тревожный вызов.Хотя первоначальная эволюция вызова тревоги у этих видов, возможно, потребовала эволюции модели дневной активности, вызов тревоги впоследствии может поддерживаться текущей адаптивной полезностью социальных, семейных выгод.
Нефилогенетический анализ и филогенетический тест концентрированных изменений указывают на связь между социальностью и суточностью, но они специально не проверяют направленность. Однако тест условных состояний показывает, что суточная жизнь не привела напрямую к эволюции социальности.Это открытие устраняет потенциальный косвенный путь развития тревожных звонков; Виды, развившие дневной образ жизни, не с большей вероятностью эволюционировали в социальную жизнь, а затем, впоследствии, вызвали тревогу. Напротив, эволюция суточного режима, по-видимому, является преимущественно и независимой причиной эволюции сигналов тревоги у грызунов. Таким образом, хотя есть два возможных пути эволюции сигнала тревоги, что удивительно, учитывая важность его адаптивной полезности, социальность может иметь второстепенное значение для грызунов.
Андерсон С., Вудс, Калифорния, Морган Г.С., Оливер В.Л.Р., 1983. Geocapromys brownii . Mammal Sp 201: 1–5.
Best TL, 1988. Dipodomys spectabilis . Mammal Sp 311: 1–10.
Best TL, 1991. Dipodomys nitratoides . Mammal Sp 381: 1–7.
Best TL, 1995. Sciurus deppei . Mammal Sp 505: 1–5.
Best TL, 1995. Sciurus variegatoides . Mammal Sp 500: 1–6.
Best TL, 1995. Spermophilus adocetus . Mammal Sp 504: 1–4.
Best TL, 1995. Spermophilus mohavensis . Mammal Sp 509: 1–7.
Best TL, Burt SL, Bartig JL, 1994. Tamias quadrivittatus . Mammal Sp 466: 1–7.
Best TL, Granai NJ, 1994. Tamias obscurus . Mammal Sp 472: 1–6.
Best TL, Hildreth NJ, Jones C, 1989. Dipodomys deserti .Mammal Sp 339: 1–8.
Best TL, Skupski MP, 1994. Perognathus flavus . Mammal Sp 471: 1–10.
Best TL, Titus AS, Caesar K, Lewis CL, 1990. Ammospermophilus harrisii . Mammal Sp 366: 1–7.
Mammal Sp 366: 1–7.
Best TL, Titus AS, Lewis CL, Caesar K, 1990. Ammospermophilus nelsoni . Mammal Sp 367: 1–7.
Биркенхольц, Германия, 1972 г. Neofiber alleni . Мамм Сп 15: 1–4.
Branch LC, Villareal D, Fowler GS, 1994.Факторы, влияющие на динамику популяций равнинной вискачи ( Lagostomus maximus , Mammalia, Chinchillidae) в зарослях кустарников в центральной части Аргентины. J Zool Lond 232: 383–395.
Brand LR, 1976. Вокальный репертуар бурундуков (род Eutamias ) в Калифорнии. Анимационное поведение 24: 319–335.
Браун Дж. К., Марес Массачусетс, 1989. Neotoma micropus . Mammal Sp 330: 1–9.
Броннер Дж., Гордон С., Мистер Дж., 1988. Отомис ирроратус . Mammal Sp 308: 1–6.
Брукс Р.Дж., Бэнкс Е.М., 1973. Поведенческая биология лемминга колларда ( Dicrostonyx groenlandians [Trail]): анализ акустической коммуникации. Anim Behav Monogr 6: 1–83.
Брудзинский С.М., 2001. Фармакологические и поведенческие характеристики сигналов тревоги 22 кГц у крыс. Neurosci Biobehav Rev 25: 611–617.
Campos CM, Tognelli MF, Ojeda RA, 2001. Dolichotis patagonum .Mammal Sp 652: 1–5.
Carraway LN, Verts BJ, 1991. Neotoma fuscipes . Mammal Sp 386: 1–10.
Carraway LN, Verts BJ, 1993. Aplodontia rufa . Mammal Sp 431: 1–10.
Carraway LN, Verts BJ, 1994. Sciurus griseus . Mammal Sp 474: 1–7.
Ceballos-G. G, Wilson DE, 1985. Cynomys mexicanus . Mammal Sp 248: 1–3.
Сервантес Ф.А., Соса В.Дж., Мартинес Дж., Гонсалес Р.М., Даулер Р.С., 1993. Pappogeomys tylorhinus . Mammal Sp 433: 1–4.
Чапман Дж. А., Фельдхамер Г. А., 1982. Дикие млекопитающие Северной Америки: биология, менеджмент и экономика.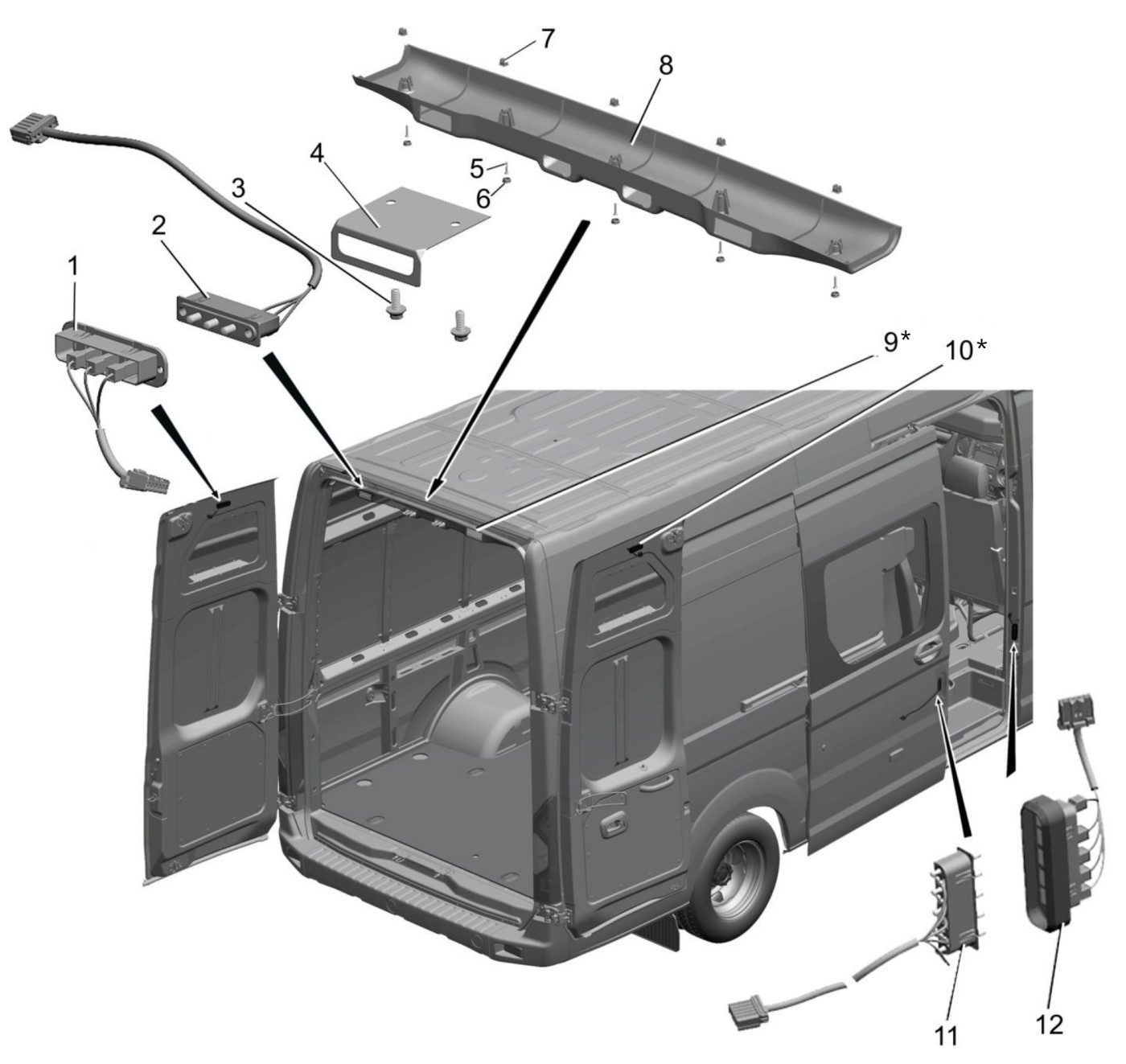 Балтимор, Мэриленд: издательство Университета Джона Хопкинса; 1147.
Балтимор, Мэриленд: издательство Университета Джона Хопкинса; 1147.
Clark TW, Hoffmann RS, Nadler CF, 1971. Cynomys leucurus . Mammal Sp 7: 1–4.
Clawson RG, Clawson JA, Best TL, 1994. Tamias alpinus . Mammal Sp 461: 1–6.
Clawson RG, Clawson JA, Best TL, 1994. Тамиас quadrimaculatus . Mammal Sp 469: 1–6.
Conroy CJ, Cook JA, 1999. Microtus xanthognathus . Mammal Sp 627: 1–5.
Cornely JE, Baker RJ, 1986. Neotoma mexicana . Mammal Sp 262: 1–7.
Дейли М., Дейли С., 1975. Социоэкология сахарских песчанок, особенно Meriones libycus . Mammalia 39: 289–311.
Давидов-Генри Б. Р., Джонс Дж. К. Дж., Холландер Р. Р., 1989. Cratogeomys castanops .Mammal Sp 338: 1–6.
de Graaff G, 1981. Грызуны Южной Африки: заметки об их идентификации, распространении, экологии и таксономии. Дурбан: Баттервортс.
Diaz GB, Ojeda RA, Gallardo MH, Giannoni SM, 2000. Tympanoctomys barrerae . Mammal Sp 646: 1–4.
Долан П.Г., Картер, округ Колумбия, 1977 г. Glaucomys volans . Mammal Sp 78: 1–6.
Эбенспергер Л.А., 1998. Социальность у грызунов: фоссориальные гистрикогнаты Нового Света как модели исследования.Преподобный Чил Хист Нат 71: 65–77.
Eisenberg JF, 1974. Функция и мотивационная основа вокализации гистрикоморфа. Symp Zool Soc London 34: 211–247.
Эйзенберг Дж. Ф., 1989. Млекопитающие неотропов, Vol. 1: северные неотропы: Панама, Колумбия, Венесуэла, Гайана, Суринам, Французская Гвиана. Чикаго: Издательство Чикагского университета.
Eisenberg JF, Redford KH, 1999. Млекопитающие неотропов, Vol. 3: центральные неотропы: Эквадор, Перу, Боливия, Бразилия.Чикаго: Издательство Чикагского университета.
Elliott CL, Flinders JT, 1991. Spermophilus columbianus . Mammal Sp 372: 1–9.
Spermophilus columbianus . Mammal Sp 372: 1–9.
Эммонс Л.Х., 1978. Звуковое общение между африканскими белками тропических лесов. Z Tierpsychol 47: 1–49.
Эрнест К.А., Марес Массачусетс, 1987. Spermophilus tereticaudus . Mammal Sp 274: 1–9.
Eshelman BD, Cameron GN, 1987. Baiomys taylori . Mammal Sp 285: 1–7.
Эшельман Б.Д., Зоннеманн С.С., 2000. Spermophilus armatus . Mammal Sp 637: 1–6.
Francescoli G, 2001. Голосовые сигналы от Ctenomys pearsoni детенышей. Acta Theriol 46: 327–330.
Francescoli G, 2002. Географические вариации голосовых сигналов Ctenomys pearsoni . Acta Theriol 47: 35–44.
Frase BA, Hoffmann RS, 1980. Marmota flaviventris . Mammal Sp 135: 1–8.
Fulk GW, 1976. Заметки об активности, размножении и социальном поведении Octodon degus .J Mammal 57: 495–505.
Gannon WL, 1988. Zapus trinotatus . Mammal Sp 315: 1–5.
Gannon WL, Lawlor TE, 1989. Вариация чип-вокализации трех видов таунсендских бурундуков (род Eutamias ). J Mammal 70: 740–753.
Garrison TE, Best TL, 1990. Dipodomys ordii . Mammal Sp 353: 1–10.
Gharaibeh BM, Jones C., 1996. Myosciurus pumilio . Mammal Sp 523: 1–3.
Джакалоне Дж., Уэллс Н., Уиллис Дж., 1987. Наблюдения за Syntheosciurus brochus (Sciuridae) в национальном парке Вулкан Поас, Коста-Рика. J Mammal 68: 145–147.
Gulotta EF, 1971. Meriones unguiculatus . Mammal Sp 3: 1–5.
Халтенорт Т., Диллер Х., 1980. Полевой справочник по млекопитающим Африки, включая Мадагаскар. Лондон: Коллинз.
Hayssen V, 1991. Dipodomys microps .Mammal Sp 389: 1–9.
Hodgdon HE, Larson JS, 1973. Некоторые половые различия в поведении в колонии меченых бобров ( Castor canadensis ). Анимационное поведение 21: 147–152.
Некоторые половые различия в поведении в колонии меченых бобров ( Castor canadensis ). Анимационное поведение 21: 147–152.
Хоффмайстер Д.Ф., 1986. Млекопитающие Аризоны . Тусон, Аризона: Университет Аризоны Press.
Hoogland JL, 1996. Cynomys ludovicianus . Mammal Sp 535: 1–10.
Hudson WH, 1872. О повадках вискачи ( Lagostomus trichodactylus ).Proc Zool Soc Lond 1872: 822–833.
Intress C, Best TL, 1990. Dipodomys panamintinus . Mammal Sp 354: 1–7.
Jackson JE, Branch LC, Villarreal D, 1996. Lagostomus maximus . Mammal Sp 543: 1–6.
Jenkins SH, Busher PE, 1979. Castor canadensis . Mammal Sp 120: 1–8.
Jenkins SH, Eshelman BD, 1984. Spermophilus beldingi . Mammal Sp 221: 1–8.
Джонсон Д.В., Армстронг Д.М., 1987. Peromyscus crinitus . Mammal Sp 287: 1–8.
Kelt DA, 1988. Dipodomys heermanni . Mammal Sp 323: 1–7.
Kingdon J, 1974. Восточноафриканские млекопитающие: атлас эволюции в Африке: зайцы и грызуны. Нью-Йорк: Academic Press.
Клейман Д.Г., 1974. Модели поведения у грызунов-гистрикоморф. Symp Zool Soc Lond 34: 171–209.
Koffler BR, 1972. Meriones crassus . Mammal Sp 9: 1–4.
Koprowski JL, 1994. Sciurus niger . Mammal Sp 479: 1–9.
Квечински Г.Г., 1998. Marmota monax . Mammal Sp 591: 1–8.
Lacher TE, Jr., 1981. Сравнительное социальное поведение Kerodon rupestris и Galea spixii и эволюция поведения у Caviidae. Bull Carnegie Mus Nat Hist 17: 1–71.
Linzey DW, Packard RL, 1977. Ochrotomys nuttalli .Mammal Sp 75: 1–6.
Макдональд Д., 1984. Энциклопедия млекопитающих. Нью-Йорк: факты в файле; 895.
McCarty R, 1975. Onychomys torridus . Mammal Sp 59: 1–5.
Onychomys torridus . Mammal Sp 59: 1–5.
McCarty R, 1978. Onychomys leucogaster . Mammal Sp 87: 1–6.
McGhee ME, Genoways HH, 1978. Liomys pictus . Mammal Sp 83: 1–5.
Medway L, 1978. Дикие млекопитающие Малайи (полуостров Малайзия) и Сингапура, 2-е изд.Куала-Лумпур: Издательство Оксфордского университета.
Merritt JF, 1978. Peromyscus californicus . Mammal Sp 85: 1–6.
Michener GR, Koeppl JW, 1985. Spermophilus richardsonii . Mammal Sp 243: 1–8.
Mones A, Ojasti J, 1986. Hydrochoerus hydrochaeris . Mammal Sp 264: 1–7.
Nagorsen DW, 1987. Marmota vancouverensis . Mammal Sp 270: 1–5.
Нэш Д. Д., Матрос Р. Н., 1977. Sciurus aberti . Mammal Sp 80: 1–5.
Никольский А.А., Суханова М.В., 1992. Ситуационно-зависимые варианты крика большой песчанки Rhombomys optimus и бегущей в норы полевки Брандта Microtus brandti . Зоол Ж 71: 125–132.
Нитикман Л.З., 1985. Sciurus granatensis . Mammal Sp 246: 1–8.
Новак Р.М., 1999. Млекопитающие Уокера в мире, 6-е изд. Балтимор, Мэриленд: Издательство Университета Джона Хопкинса.
Новак Р. М., Парадизо Дж. Л., 1983. Млекопитающие Уокера, 4-е изд. Балтимор, Мэриленд: Издательство Университета Джона Хопкинса.
Oaks EC, Young PJ, Kirkland GL Jr, Schmidt DF, 1987. Spermophilus variegatus . Mammal Sp 272: 1–8.
O’Farrell MJ, Blaustein AR, 1974. Microdipodops megacephalus . Mammal Sp 46: 1–3.
O’Shea TJ, 1991. Xerus rutilus . Mammal Sp 370: 1–5.
Pardinas UFJ, Galliari CA, 2001. Ритродон ауритус . Mammal Sp 664: 1–8.
Полсон Д.Д., 1988. Chaetodipus hispidus . Mammal Sp 320: 1–4.
Пеппер Дж. У., Барауд Ш., Лейси Э. А., Шерман П. У., 1991. Вокализации голого землекопа. В: Биология голого землекопа (Sherman PW, Jarvis JUM, Alexander RD, ред.). Принстон, Нью-Джерси: Издательство Принстонского университета; 243–274.
Perez EM, 1992. Agouti paca . Mammal Sp 404: 1–7.
Perrin MR, Dempster ER, Downs CT, 1999. Gerbillurus paeba . Mammal Sp 606: 1–6.
Pessoa LM, dos Reis SF, 1993. Proechimys dimidiatus . Mammal Sp 441: 1–3.
Pessoa LM, dos Reis SF, 1996. Proechimys iheringi . Mammal Sp 536: 1–4.
Pessoa LM, dos Reis SF, 2002. Proechimys albispinus . Млекопитающие Sp 693: 1–3.
Pizzimenti JJ, Collier GD, 1975. Cynomys parvidens . Mammal Sp 52: 1–3.
Pizzimenti JJ, Hoffmann RS, 1973. Cynomys gunnisoni . Mammal Sp 25: 1–4.
Randall JA, 1994. Конвергенции и расхождения в коммуникации и социальной организации пустынных грызунов. Aust J Zool 42: 405–433.
Рэндалл Дж. А., 2001. Эволюция и функция барабанной дроби как коммуникации у млекопитающих. Am Zool 41: 1143–1156.
Рэндалл Дж. А., Роговин К. А., Шиер Д. М., 2000. Антихищническое поведение социальных грызунов пустыни: барабанная дробь и тревожный вызов у большой песчанки, Rhombomys opiums .Behav Ecol Sociobiol 48: 110–118.
Редфорд К. Х., Эйзенберг Дж. Ф., 1992. Млекопитающие неотропов, Vol. 2: южный конус. Чикаго: Издательство Чикагского университета.
Рейх Л.М., 1981. Microtus pennsylvanicus . Mammal Sp 159: 1–8.
Рейг О.А., 1970. Экологические заметки окаменелого грызуна-октодонта Spalacopus cyanus (Molina). J Mammal 51: 592–601.
Rickart EA, 1987. Spermophilus townsendii .Mammal Sp 268: 1–6.
Рикарт Э. А., Йенсен Э., 1991. Spermophilus Washingtoni . Mammal Sp 371: 1–5.
А., Йенсен Э., 1991. Spermophilus Washingtoni . Mammal Sp 371: 1–5.
Робертс Т.Дж., 1977 г. Млекопитающие Пакистана. Лондон: Ernest Benn Ltd.
Rood JP, 1972. Экологические и поведенческие сравнения трех родов аргентинских морских свинок. Anim Behav Monogr 5: 1–83.
Schleich CE, Busch C, 2002. Ювенильные вокализации Ctenomys talarum (Rodentia: Octodontidae).Acta Theriol 47: 25–33.
Зигель Х.И., 1985. Хомяк: размножение и поведение. В. Нью-Йорк: Пленум Пресс; 440.
Смолен М.Дж., 1981. Microtus pinetorum . Mammal Sp 147: 1–7.
Снайдер Д.П., 1982 г. Tamias striatus . Mammal Sp 168: 1–8.
Соколов В.Е., Котенкова Е.В., Микаиленко А.Г., 1998. Mus spicilegus . Mammal Sp 592: 1–6.
Стойл ДТ, 1990. Microtus ochrogaster .Mammal Sp 355: 1–9.
Стил М.А., 1998. Tamiasciurus hudsonicus . Mammal Sp 586: 1–9.
Стил М.А., 1999. Tamiasciurus douglasii . Mammal Sp 630: 1–8.
Streubel DP, Fitzgerald JP, 1978. Spermophilus tridecemlineatus . Mammal Sp 103: 1–5.
Стуббе А., Янке С., 1994. Некоторые аспекты социального поведения у полевок Microtus brandti (Radde, 1861). Pol Ecol Stud 20: 449–457.
Sutton DA, 1992. Tamias amoenus . Mammal Sp 390: 1–8.
Тамура Н., Хаяси Ф., Мияшита К., 1988. Иерархия доминирования и поведение при спаривании формозской белки Callosciurus erythraeus thaiwanensis . J Mammal 69: 320–331.
Тоннелли М.Ф., Кампос С.М., Охеда, РА, 2001. Microcavia australis . Mammal Sp 648: 1–4.
Torres-Mura JC, Contreras L, 1998. Spalacopus cyanus .Mammal Sp 594: 1–5.
Van Den Brink FH, 1968. Полевой справочник по млекопитающим Великобритании и Европы. Бостон: Компания Houghton Mifflin.
Бостон: Компания Houghton Mifflin.
Veal R, Caire W, 1980. Peromyscus eremicus . Mammal Sp 118: 1–6.
Вейтл С., Бегалл С., Бурда Х, 2000. Экологические детерминанты параметров вокализации: случай Coruro Spalacopus cyanus (Octodontidae), ископаемого социального грызуна. Биоакустика 11: 129–148.
Verts BJ, Carraway LN, 1987. Thomomys bulbivorus . Mammal Sp 273: 1–4.
Verts BJ, Carraway LN, 2002. Neotoma lepida . Mammal Sp 699: 1–12.
Verts BJ, Kirkland GL, Jr., 1988. Perognathus parvus . Mammal Sp 318: 1–8.
Вт CHS, 1975. Вокализация австралийских прыгающих мышей (Rodentia, Notomys ). Дж. Зоол 177: 247–263.
Wells NM, Giacalone J, 1985. Syntheosciurus brochus .Mammal Sp 249: 1–3.
Wells-Gosling N, Heaney LR, 1984. Glaucomys sabrinus . Mammal Sp 229: 1–8.
Whitaker JO, JR, 1972. Zapus hudsonius . Mammal Sp 11: 1–7.
Whitaker JO Jr, Wrigley RE, 1972. Napaeozapus insignis . Mammal Sp 14: 1–6.
Whitaker JO Jr, 1996. Полевой справочник по североамериканским млекопитающим Национального общества Одубона. Нью-Йорк: Кнопф.
White TG, Albercio MS, 1992. Dinomys branickii . Mammal Sp 410: 1–5.
Wiley RW, 1980. Neotoma floridana . Mammal Sp 139: 1–7.
Williams DF, Kilburn KS, 1991. Dipodomys ingens . Mammal Sp 377: 1–7.
Williams LR, Cameron GN, 1991. Geomys attwateri . Mammal Sp 382: 1–5.
Williams SL, 1982 г. Geomys personatus . Mammal Sp 170: 1–5.
Виллнер Г.Р., Фельдхамер Г.А., Цукер Е.Е., Чепмен Дж. А., 1980. Ondatra zibethicus . Mammal Sp 141: 1–8.
Wilson DE, Ruff S, 1999. Смитсоновская книга североамериканских млекопитающих. Вашингтон, округ Колумбия: Пресса Смитсоновского института; 750.
Вашингтон, округ Колумбия: Пресса Смитсоновского института; 750.
Woods CA, 1973. Erethizon dorsatum . Mammal Sp 29: 1–6.
Woods CA, Boraker DK, 1975. Octodon degus . Mammal Sp 67: 1–5.
Woods CA, Contreras L, Willner-Chapman G, Whidden HP, 1992. Myocastor coypus .Mammal Sp 398: 1–8.
Йенсен Э., Шерман П.В., 1997. Spermophilus brunneus . Mammal Sp 560: 1–5.
Young CJ, Jones JK, Jr, 1982. Spermophilus mexicanus . Mammal Sp 164: 1–4.
Янгмен П.М., 1975 год. Млекопитающие территории Юкон. Оттава: Национальный музей Канады.
Зегерс Д.А., 1984. Spermophilus elegans . Mammal Sp 214: 1–7.
Благодарим T. Best, E. Heymann, C.Knogge и M. Mares за экспертную помощь с недостающими данными; Дж. Хэйр, П. Наринс, П. Нонакс, Т. Орд, К. Поллард, М. Зук и анонимный рецензент за комментарии к предыдущим версиям рукописи; и Отделение естественных наук Калифорнийского университета в Лос-Анджелесе за частичную поддержку посредством создания фондов для D.T.B.
Поведенческая экология, том. 16 нет. 1 © Международное общество поведенческой экологии, 2005; все права защищены.
Ложные тревоги: 7 причин, почему они у нас есть
Беспокойство естественно.
Спокойствие познается.
Страх помог нашим предкам выжить в опасном мире, поэтому мы унаследовали ум, который хорошо переносит страх. Он выделяет опасные химические вещества, даже если вы действительно в безопасности.
Вот семь причин, по которым наши опасные химические вещества дают ложные срабатывания. Когда вы поймете свой мозг, вы сможете изменить его в более счастливом направлении.
№1. Беспокойство — одно из самых первых переживаний.
Кортизол — это химическое вещество, которое выделяется, когда вы видите угрозу для выживания. Вы родились с чувством угрозы выживанию, потому что были голодны и замерзли, и у вас не было возможности удовлетворить свои потребности. Кортизол заставлял вас плакать, и это помогало вам выжить. Со временем вы научились удовлетворять свои собственные потребности в выживании, но когда вы думаете, что не можете этого сделать, уровень кортизола резко возрастает.
Вы родились с чувством угрозы выживанию, потому что были голодны и замерзли, и у вас не было возможности удовлетворить свои потребности. Кортизол заставлял вас плакать, и это помогало вам выжить. Со временем вы научились удовлетворять свои собственные потребности в выживании, но когда вы думаете, что не можете этого сделать, уровень кортизола резко возрастает.
№2. Нейроны соединяются, когда течет кортизол.
Когда вы чувствуете угрозу, кортизол помогает проложить путь, который быстрее вызовет плохие чувства, когда вы в следующий раз увидите что-то подобное.Это защитит вас от двойного прикосновения к горячей плите, но также может заставить вас чувствовать себя плохо из-за вещей, которые вам нравятся, например, учебы или социальных инициатив.
№ 3. Кортизол привлекает ваше внимание.
Кортизол заставляет вас чувствовать себя так плохо, что вы не можете ни о чем думать, кроме как заставить его остановиться. Кортизол побуждает газелей убегать от хищников, когда они предпочитают продолжать есть. Кортизол разработан, чтобы помочь вам убежать от вреда, поэтому он вызывает у всего тела чувство тревоги.
№4. Кортизол заставляет ваш мозг сканировать на наличие угроз.
Когда газель нюхает льва, как она узнает, куда бежать? Кортизол побуждает мозг находить подробности о потенциальных угрозах. Ваш большой мозг умеет находить сигналы угрозы, когда он смотрит.
№ 5. Наш мозг учится на наградах.
Беспокойство иногда приносит вознаграждение, например социальную поддержку или признание. Ваши счастливые химические вещества текут и побуждают вас повторять поведение, которое было вознаграждено.Вы можете повторять тревожные мысли из-за этого положительного ожидания.
№ 6. Разочарование может вызвать выработку кортизола.
Вы сознательно не воспринимаете разочарование как угрозу выживанию, но по своей природе неудача означает голод. Кортизол говорит льву, чтобы он прекратил тратить энергию на сбежавшую газель. Ваш кортизол может встревожить вас, когда ваши усилия не увенчаются успехом, даже если вы не собираетесь голодать.
Кортизол говорит льву, чтобы он прекратил тратить энергию на сбежавшую газель. Ваш кортизол может встревожить вас, когда ваши усилия не увенчаются успехом, даже если вы не собираетесь голодать.
№ 7. Расстройство статуса также может вызвать выработку кортизола.
Мы часто не признаем, что нас волнует статус, но животные соревнуются за статус, как будто их жизнь зависит от него, потому что это помогает увековечить их гены. Мы унаследовали мозг, который может выделять кортизол, когда мы видим потенциальную угрозу нашему социальному статусу.
Вы можете построить в своем мозгу новые пути, чтобы чувствовать себя в безопасности в мире таким, какой он есть. Забавный и быстрый способ сделать это можно найти в моей книге T ame Your Anxiety: Reewiring Your Brain for Happiness .
Друзья Детской фермы Парк
Программа животноводства Общества друзей Детской фермы-парка
РАСПИСАНИЕ РОЖДЕНИЙ НА 2019 ГОД
Коровы
Сникерс
Свиньи
Лилия 4Н
Молочные козы
Василий
Отряд коз
Пенни
Младенцы Пенни
Эдит
Младенец Эдит
Начальная стадия нарушения баланса питательных веществ в организме.
Что такое тревожная стадия стресса?
Стадия тревоги стресса — это начальная реакция на стресс , обычно называемая стадией стресса «бей или беги» .Тревожная стадия стресса — это вегетативная реакция, производимая осью HPA, предназначенная для выживания и инициирующая огромную потребность в питательных веществах в вашем теле. Тревожная стадия стресса может быть вызвана физической угрозой изнутри, такой как бактериальная или вирусная атака, или извне, предполагаемой или реальной психологической угрозой или физической атакой.
Ваши эмоции также влияют на вашу реакцию на стресс и должны учитываться в Управление стрессом . Негативные эмоции, такие как ненависть, зависть, страх, печаль и гнев, могут вызывать нездоровую индивидуальную реакцию на стресс.Частота и продолжительность отрицательных эмоций могут легко отрицательно повлиять на ваш статус питания.
Тревожная стадия стресса мгновенно вызывает психофизиологические изменения. Ваша симпатическая нейроэндокринная система вступает во владение мгновенно; ваше тело производит поток мощных гормонов стресса (например, адреналина). Теперь вы находитесь в режиме «сражайся или беги».
Ваши зрачки расширяются; ваша пищеварительная система замедляется, частота сердечных сокращений повышается, кровяное давление повышается, уровень глюкозы повышается, ваша сила увеличивается, и происходит множество других симпатических нейроэндокринных изменений.Все эти изменения предъявляют огромные требования к вашей биохимии питания.
Тревожная стадия стресса провоспалительная из-за избыточной выработки альдостерона. Пребывание в стадии тревоги вызывает хроническое воспаление и может привести к множеству симптомов или проявлений .
Пребывание в стадии тревоги вызывает хроническое воспаление и может привести к множеству симптомов или проявлений .
Анализ волос и тревожная стадия стресса
Анализ волос может выявить тревожную стадию стресса. Тем не менее, для правильной интерпретации важно учитывать многие закономерности, выявленные в результатах.Сюда входят токсичных элементов . На графике результатов вы увидите повышенный уровень натрия по сравнению с калием. Натрий, стимулирующее питательное вещество, увеличивается за счет удерживания через сигнальную систему от оси HPA к надпочечникам, которая сигнализирует почкам о необходимости удержания натрия.
По мере увеличения содержания натрия он становится антагонистическим по отношению к магнию (и многим другим питательным веществам) и начинает создавать дефицит. Магний — это буфер для стресса и успокаивающее питательное вещество. Это увеличивает соотношение натрий / магний и снижает соотношение кальций / калий.
Другие минеральные питательные вещества, благодаря усиленному удерживанию, использованию или выведению, включают калий, цинк, медь, кобальт, хром и селен. Это одновременно стимулирующие и седативные питательные вещества.
Неполный список дополнительных питательных веществ включает витамины C, D, E, B1, B6 и B12. Из-за динамики взаимосвязей питательных веществ, когда каждое важное питательное вещество зависит от другого (кофакторов), они, в свою очередь, влияют на все аминокислоты.
Например, изолейцин, лейцин и валин являются аминокислотами с разветвленной цепью (BCAA) и больше связаны с физической стороной стресса.Метаболически им необходимы витамины B6 и B1. Затем им в качестве кофакторов требуются производные меди и B2. Наконец, для правильного метаболизма BCAA необходимы биотин, магний и альфа-кетоглутарат, полученные из глутаминовой кислоты. Нарушение прямых и косвенных взаимосвязей между питательными веществами происходит на всех стадиях стресса.
Лизин, тирозин, фенилаланин, цистеин, цитруллин и аспарагиновая кислота — это аминокислоты, связанные с умственным или психологическим стрессом. Метаболические функции этих аминокислот дополнительно нарушаются из-за требований, предъявляемых к их витаминным и минеральным кофакторам во время стресса.
Находитесь ли вы в тревожной стадии стресса? Закажите анализ волос сегодня.
Следуй за Газелью!Nature является прекрасным примером того, как «должна» разворачиваться реакция на стресс. Вы можете следить за газелью на всех этапах стресса. Мы выбрали газель, потому что думаем, что большинство людей видели шоу природы с участием газели у водопоя.
Когда газель находится у водопоя и пьет немного воды, аллигатор внезапно бросается к газели.Мгновенно газель переживает тревожную стадию стресса. Это известно как стадия стресса «борись или беги». В нашей газели задействованы все системы выживания, и она пытается убежать. В случае успеха наша газель перейдет в стадию сопротивления стрессу .
Делитесь знаниями!
Если вы нашли эту страницу информативной и полезной, поделитесь ею со своей семьей и друзьями. Если вы выберете HairAnalysisReport.com в качестве провайдера, поделитесь и этим!
Содержимое и лабораторные услуги, представленные на этом сайте, предназначены только для образовательных и информационных целей и не предназначены для диагностики, лечения, предотвращения или лечения заболеваний.
Ссылки на стресс
.